
The Specificity of Words & Nerves | 3
What the nervous system reveals about sensation, language & communication:

Neurolinguistic pathways are localised yet connected:
The first distinction we make now in this article series on language, is that our senses provide us with information but not with comprehension or interpretation of it.
Neither do our senses prioritise certain aspects of information over others, although they may be sensitised or change modes, such as during the adjustment of the human ear to low volume sounds, or of the eye to darkness.
This distinction shows that the human nervous system is fundamentally wired for the adaptive gathering of information prior to the prioritisation of certain objectives over others, whether or not we are consciously aware of these objectives. Our brains for example, know we are thirsty or fatigued before we become consciously aware of the process.
Like the cerebral functions of linguistic comprehension and interpretation, the general somatic sensations discussed in the previous article are a function of the brain itself, not of the tissue which comes into contact with the stimulus. When the hand is dipped in cold water, the sensation is decodified by the brain, not by the peripheral nerves which codify cold and wetness.
Suffice it to say, linguistic specificity is dependent on the comprehension, interpretation, and fluency of language. As such the codification of language must be learned, but the human brain nonetheless has a neurological predisposition for it.
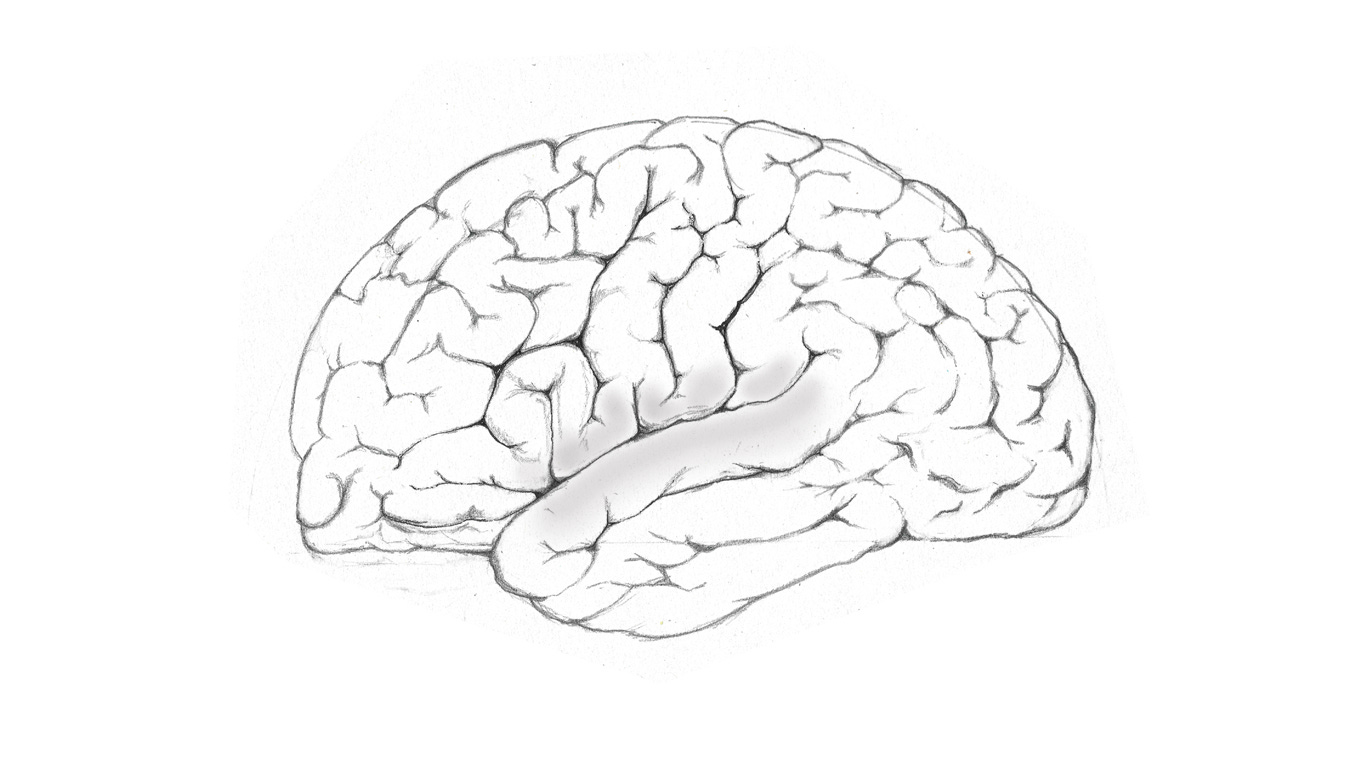
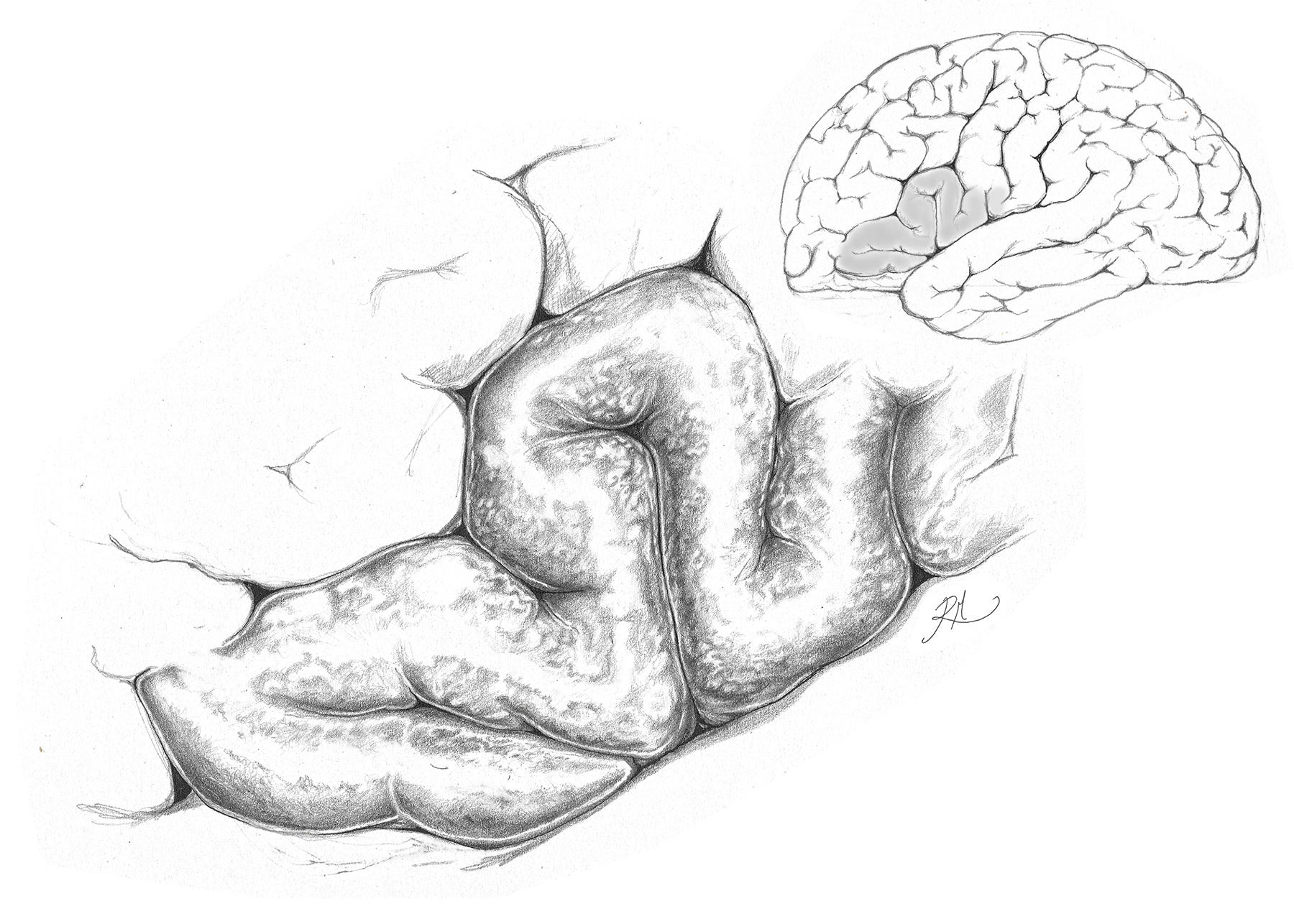
Linguistic functions are localised to the temporoparietal region of the cerebrum shown above, where the temporal and parietal lobes abut each other, and are separated by an oblique sulcus (a narrow groove), known as the fissure of Sylvius.
The neural function of the gyri that are responsible for language, are topographically labelled the inferior frontal and the superior temporal gyri, but are functionally known as Broca’s and Wernicke’s areas respectively. They serve as adjunct parts of the cerebrum, that outer tissue which makes up the majority of our brain, and among many functions provides us with the capacity to organise and integrate our conscious thoughts, to exercise reasoning and judgement, engage in learning, problem solving, and much else besides.
These latter functions are most directly the remit of the frontal lobes, but require interconnectivity with other brain regions to operate at an optimum. This includes language, speech, articulation, comprehension, interpretation and audition, which are distinct yet continuous in terms of their neuroanatomical functions during human communication.
The inferior medial gyrus is responsible for verbal expression, and contains Broca’s area. It forms part of the cortices, the ‘outer bark’, as below.
Visual sections through the brain, as seen on an imaging scan, are referred to as planes of section. When looking at any plane of section through the brain, it becomes evident that a significant proportion of the cerebrum is dedicated to the white matter fibres which fan upward into the outermost layer of grey matter. These fibres are responsible for mediating neural transmissions which originate in the peripheral nervous system, the brainstem nuclei or contralateral hemisphere.
This white matter makes up approximately half of any human cerebrum, with the remaining cerebral grey matter being distributed throughout the gyral exterior. If we consider also that the cerebrum comprises ~82% of the brain’s overall mass, we find that a vast amount of the neural real estate is dedicated to connection between modular functions within brain tissue.
As the repository of all language, the brain in and of itself has much to teach us about how and why language functions as it does, including the selection of words to solve problems of description and categorisation, and how the application of language in everyday life forms the structure of our thoughts and ideas.
The observation of neurolinguistic capacity in health relative to disease is primarily concerned with comprehension and speech production. It has led to direct clinical observations of how the function of language is influenced by change due to trauma or neurodegeneration. Through this we can gain very robust and cohesive insight into what language is objectively, and all the more intriguing is the measure of why it exists as a means of communicating with other members of our species in the first place.
We can deduce that the specificity of language in terms of words that we choose to use, and even the creation of language itself, is not just an abstraction, but rather, entails a unique dynamic between real world stimuli and physiological functions.
Recall that information may be sensed directly as a stimulus, such as light or sound, or a codification of sounds and somatic cues that we interpret and decode via language. Body language for example, is a form of somatic cue. If language ultimately represents the most highly functional means of contextualising, retaining, associating and expressing information, we might then ask, what nuances do neurolinguistic functions reveal about the human capacity for language, and language itself?
Patterns are sensed, comprehended & interpreted:
When viewed as a functioning whole, the auditory and visual systems convert audible and visible patterns of language into electrical signals, becoming a codified forms of information. This information is organised by highly configured structures within the brain which allow the transmission of the information to its appropriate place.
Tactile sensation can of course also be used to infer or to read information, whether linguistic, or as a physical stimulus which indicates a change in the environment, such as vibration or temperature. In this way, patterns from the outside world are codified into electrical transmissions. A specific pattern is made to represent a specific stimulus, and consistency between the outside world and internal response is maintained.
This consistency is crucial because the fidelity of information conveyed to the mind and body, informs the response, whether this comes from voluntary or involuntary branches of the nervous system, or indeed both. The precision with which we understand the world through the senses is a necessary precursor to our being able to describe ourselves and our situation in it.
In the event of neuropathology, neural tissues will always attempt to compensate when and wherever possible, by way of regeneration in some cases, but in cases of severe tissue damage, most often through the rerouting of the localised function to another region of tissue.
This rerouting occurs within residual tissues of the region that has been spared from damage, but may also harness the symmetry of the brain in order to compensate. The anatomical function of the brain is patently symmetrical, and as we explored in the previous part of this article series (part 2), this provides for the division of motoric and sensory functions of the nervous system into two bodily halves.
Yet not all of these brain functions are distributed in this bilateral way. With regards to language for instance, the left hemisphere is the dominant language centre among the majority of human populations.
Despite the neurolinguistic functions of the brain being dominant in the left hemisphere over the right, the two hemispheres can in certain cases, such as when the temporo-parietal area is affected by stroke, allow for damage in one hemisphere to be compensated for.
Although language networks of the brain are left hemisphere dominant in the majority of the population, this function can be reorganised into undamaged regions of the network in the ipsilateral (same side) hemisphere which have not been affected, or rerouted to the contralateral (other side) cortices of the right hemisphere.
Another compensatory mechanism which has been posited is functional reserve, which describes the recruitment of neural tissue that is unrelated to the damaged tissue in terms of function, but may nevertheless adapt to new demands placed upon it.
The brain's modular functions are numerous and their connections between each other prolific, yet this connectivity must also remain specific in order to allow for functions to occur in an appropriate way.
In processing sensory information and affecting movement to produce linguistic and lexical output for example, functions of the temporoparietal cortices are coordinated to produce forms of communication that are selective and replicable. This is yet another direct example of form preceding function.
As nerve tracts ascend and descend via grey matter into and from the brainstem, the crossing over of neural pathways from one half of the body to the other often occurs. This depends on the function of the tract and the connections it makes within the central nervous system. These innumerable connections made between nerves that innervate certain tissues are crucial to the integration of codified electrical stimuli, providing neural sensation and motor control.
The specificity of neuroanatomical terms we require in order to differentiate between common and more precise forms of neural connections are important, and the following are two principle examples:
The first is a more general term, anastomosis, which denotes an interconnection between adjacent nerve fibres, and allows a redundancy of connections to occur in the event that any are damaged. For instance if a connection between nerve A and B1 and B2 exists, then the loss of innervation to B1 can be compensated for by a connection that exists between B2 and A.
Because vascular tissues anastomose far more frequently than nerves do, the word anastomosis is more often used in relation to blood vessels. This occurs because neurobiological control over the physical dynamics of a liquid differs greatly to that of the dynamics of electric conduction.
Second is the word, decussation, which is similar in its base definition, yet is more exactly applied for example, to the crossing of connections between the hemispheres of the brain, known as the commissural nerve fibres, which travel between hemispheres through the region known as the corpus callosum.
Similarly, the optic chiasm (cross) for example, transmits the nasal (medial) half of the retinal field from one eye to the apposite (contralateral) hemisphere of the brain.
During this process the visual field is converted into electrical patterns, and is then separated into intercalated quadrants by the configuration of the fibres. Decussations also occur between the brain and the ascending and descending spinal tracts, and within the tracts themselves as they are found within the spinal cord.
Given all these interconnections, it is clear why delineating them in terms of their form has everything to do with the function of neural innervations they make. It follows that being able to specify their form is conducive to communicating their role in living tissue.
As explored previously, in some but not all cases, the crossing of nerve channels may allow for neural function to continue in the event of its loss (denervation), so that if one nerve branch is compromised, there may be a partial if not completely innervating nerve available to compensate.
When a nerve is not branched into another via anastomosis, then this nerve is said to be autonomous, and is more readily identifiable as a contributor to afferent sensation or efferent action as a result of its autonomy; this is because damage to a nerve of this kind will not be compensated for beyond the point at which it maintains lateral connection with other nerves.
Variation in how the anastomosis of a nerve occurs, if it occurs at all, can present differently in one individual when compared to the next. This is a caveat to the consistency of anatomy mentioned previously. The real world consequences of neuroanatomical variation are very real, but highly contextual. Suffice to say however, that the consistency in neuroanatomy and anatomy across populations in general, predominates because the functionality and adaptivity of the structures do.
Adapted Sensations:
At the outermost periphery of the nervous system, where the dermal layers of the skin and the muscle which lies deep to it are found, sensations may differ despite their being mediated by the same type of nerve ending.
The codification of nociception (somatic pain) for example, is evidently different to that of pruriception - which we know colloquially as ‘itching’, and yet, both sensations may be transmitted through the same nociceptive nerve endings. The stimulus is differentiated only by its codification as a pattern, sensed from the outside world, and which the nerve ending is sensitised to, and in turn which the brain interprets as itch on the one hand, or pain on the other.
We shall return to nociception as a specifier of physiologically meaningful information later. Sufficed to state the obvious here again, that your body prefers you to experience the relative discomfort of sensing what for the majority of the time is something innocuous (a false positive), than not sensing it.
This is simply because the likelihood of surviving what could be a hazard is increased. For example, surface contact with an infectious or poisonous substance (the true positive scenario), as transmissible by an insect. By brushing the insect away before you experience the pain of it damaging bodily tissues, you limit the risk of damage before it occurs.
The sensation of itch and pain being codified by the same integumentary receptor seem both an efficient and more failsafe solution. This sensation preserves the consistency between reality and what we think is happening to our bodily surfaces, by leveraging the same nerves to codify a scale of differing sensations.
It is necessary that there be both specificity and priority of neural sensation. To achieve this, the same specificity and priority may also be expressed via the language we use to differentiate between these sensations.
Up to now in this article on the specificity of language, we have used neuroanatomical science to begin to explore some key aspects of how and why all language must begin as information that has been sensed directly from physical spaces, both within the body and without.
The neuro-affective states (of emotion) that interact dynamically with our thoughts are neurophysiological, and contribute constantly to the sum of those thoughts which we might (or might not) in turn communicate with others.
Yet we still articulate our thoughts and emotions through language to ourselves when we consciously engage with them, and as such, this quiet monologue is the intermittent yet everyday language of our inner lives.
We may even reply to those emotions with unspoken reason, or vice versa, to reason with emotions that turn over constantly beneath, as if our minds were a landscape of forests, all connected and talking to each other at the same time, coordinated into a series of driven interests, a single one in waiting whilst the other is winning out.
This quite human capacity allows us to specify our thoughts within our own minds through language, and apply linguistic precision to clarify our reasoning and to define our feelings. This is especially the case when the more cognitive may be in conflict with the more affective aspects of our experience.
Whether intrinsic or extrinsic to the body, evocative or calculative, both special, general and visceral forms of neuro-sensation provide us with information about our world. Their information is collated into a meaningful understanding, which may be formulated into a concept, relatable by us to another, or by another to us.
As readers, listeners and speakers, we are evidently familiar with language, but the patterns of the language we produce in writing or speech need be decoded first by the language centres and pathways of the brain.
We shall explore this further as a combined process in the next article in this series.
Thank you for reading!