
Fibre, Root & Horn | 1:
How the architecture of the spinal cord and cranial nerves mediate our senses and shape human nature:

In this LV series, we explore…
the architecture of nerves as a mechanical means of drawing human beings towards advantage and away from danger,
the control that chemical and neuronal structures have in maintaining vital functions in everyday human life,
and how the known signs and symptoms of neuropathy are caused by abnormal alterations to these.
The cord proper…
At a diameter of only 10 to 13.5mm, the human spinal cord is widest at the neck (1). This remarkably small area houses a vast collection of nerve fibres, with approximately half their sum needed to convey sensations from the outward surface of the skin, as well as the fascia, muscles, joints and viscera which lie deep to it.
Human spinal nerves ramify through 31 levels in order to reach the tissues they innervate via electrochemical transmissions to and from the cord. With each ramified nerve projecting laterally, the width of the spinal cord diminishes as it descends from the brain. In both humans and our mammalian relatives the spinal nerves continue to ramify until they cease as the cauda equina, ‘the horse’s tail’, where the remaining nerve roots branch at the level of the sacrum, and descend into the pelvic cavity and lower limbs. All the sensations, reflexes and movements of our lower body, including the sexual organs which allow us to reproduce, are made possible through these nerve branches and the organisation of tracts within the spinal cord from which they flow. Our sense of stability in relation to the surface on which we stand is likewise a function of sensation conducted by the lower body and allows for us to orient our position, brace ourselves physically and adapt to almost any surface.
Broad comparison can be made between neural tissues at every level (whether of cells or of nerve fibres visible to the naked eye) and the function of many forms which branch in order to glean information, distribute vital fluids or gather resources. The morphology of a tree e.g. is not dissimilar to neural cells in basic shape, and the descriptive terms ‘branched’, ‘dendritic’ or ‘arborised’ have been applied to neural tissues by the earliest scientific pioneers of neuroanatomy in the 19th to 20th century, these individuals being the first to observe neural cells directly through microscopy, including Heinrich Waldeyer and Santiago Ramón y Cajal.
Such terms are still used today by anatomists and surgeons especially, as they often imply a structure relatable to the underlying functions of the tissue. A loss of this structure will result in a loss of its function, prompting the inchoate question of neurology and neuroscience at their inception: how humanity is maintained by a selective current of millivolts, controlled across multitudes of cells, signalling in sequence at the appropriate time, and ceaselessly regulating every cubic inch of organic matter in response to changes within and changes without.
Alongside sensory fibres the spinal cord contains the tracts responsible for outgoing motor signals into peripheral tissues, providing a means for both voluntary and involuntary movements, secretions, contractions and so on. Despite such limited space, vast yet highly selective signalling is made possible by the sheer organisation of nerve fibres into dedicated functions, as determined by their structure, their calibre of myelination (for electrical insulation & propagation), their situation in the body and the chemical neurotransmitters they utilise. These features dictate which tissues and other neurons will be targeted, and how electrochemical signals are codified for the brain to interpret, maintaining tone and initiating phasic responses in some instances, whilst inducing long-term transformation (such as growth) in others.
Transmissions are separated from one another much in the same way an electrical wire is insulated, where with nerves the upmost importance of this feature is in preserving the fidelity and reliability of each signal. The fibres are compartmentalised into fasciculi (bundles) and funiculi (larger bundles), with those responsible for conveying prolonged physical pain being separated from those responsible for the sensation of non-noxious stimuli, such as tactility. The loss of this compartmentalisation contributes to a variety of neurological dysfunctions which articles in this Lucida Visionis series will follow, such as hyperalgesia, allodynia and paraesthesia.
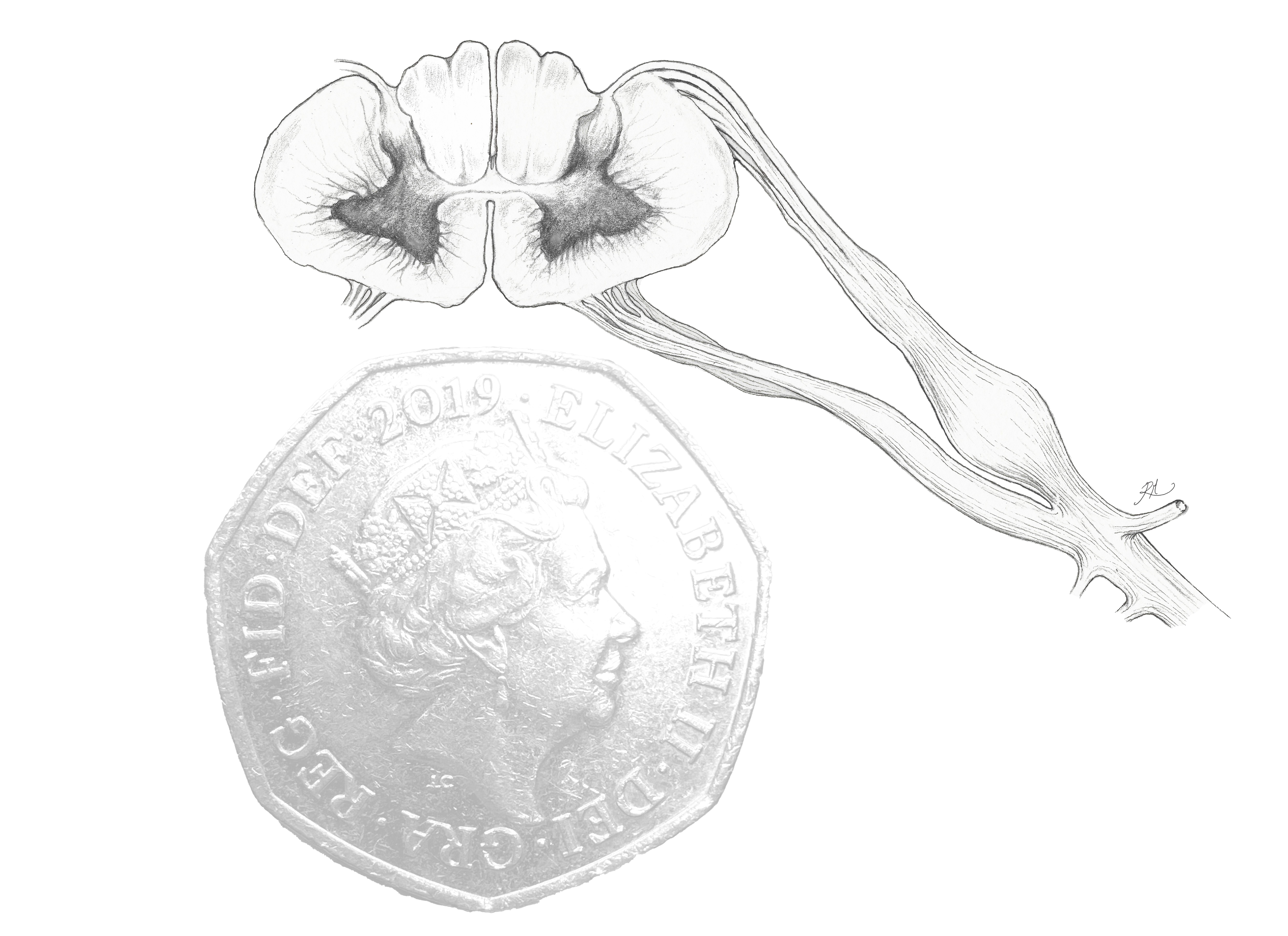
At each level of the spinal cord (the illustration above e.g., shows a cross section of the cord from the 8th cervical level) the grey matter assumes a horn like symmetry either side of the midline, comprising a dorsal (posterior) horn and a ventral (anterior) horn. The dorsal horn is populated by a majority (>90%) of dense interneuronal networks. The arborised axons of these neurons are restricted to the dorsal horn, where they serve to promote or inhibit sensory transmissions to the brain, limiting or enhancing the long distance transmissions conveyed by 1st order projection neurons which target tissues peripheral to the cord (via the dorsal root ganglion shown) and 2nd order projection neurons that project from within the cord to the brain. These include those signals from free nerve endings which convey painful or pleasurable sensations, or general tactile, follicular (hair) and thermal sensations from bodily surfaces, being at their most distal and numerous from the brain in the glabrous skin that is found overlying the palm of the hand, including the skin overlying the palmar aspect of the fingers. This applies also to the soles of the feet.
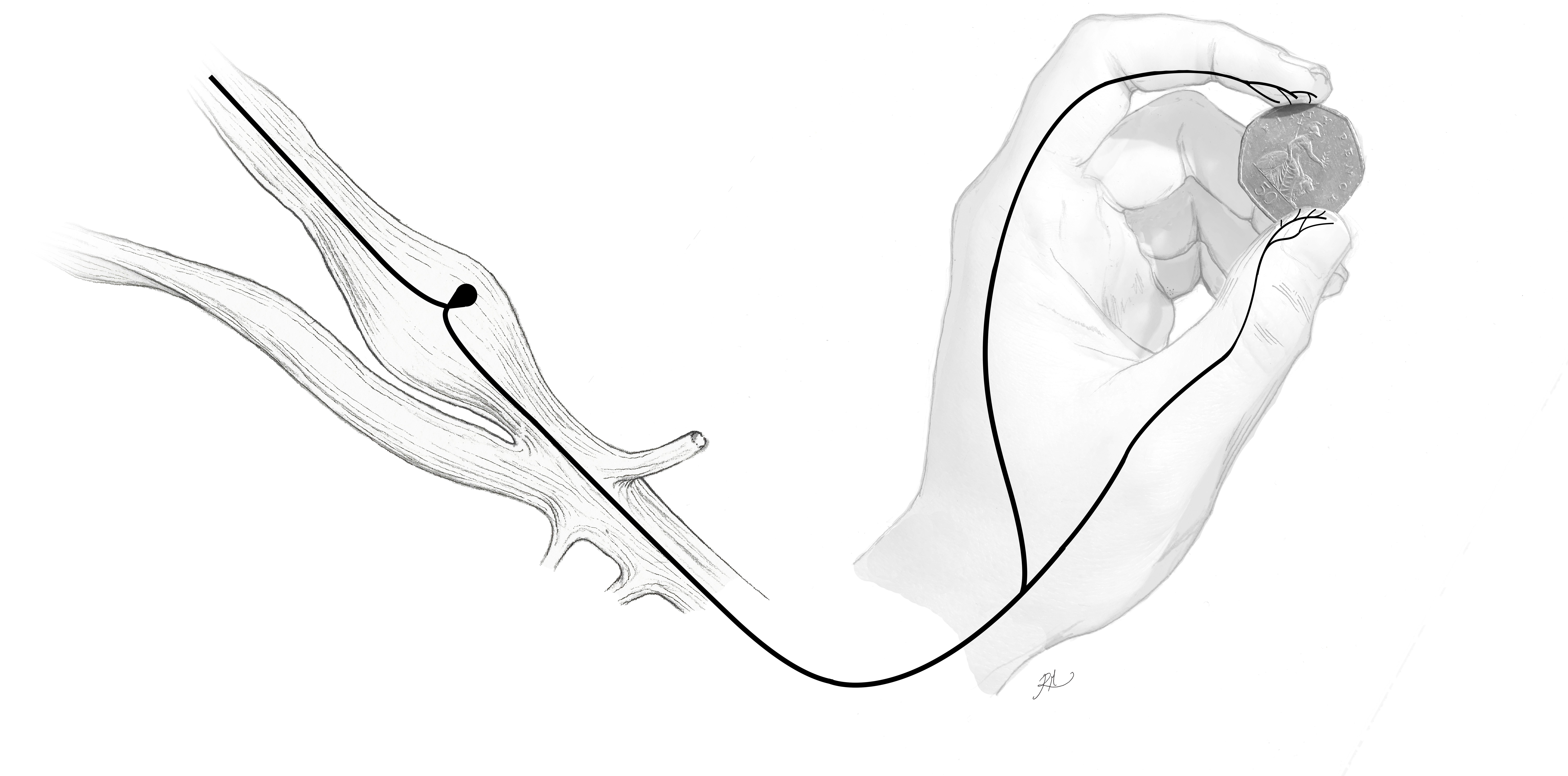
Human dexterity allows us the use of endlessly varied tools, and to experience uniquely diverse sensations, both in response to our environment and in social activities. The importance of context in allowing us to define all these neurosensory signals as we experience them cannot be understated. This is itself achieved through separation of the sensory fibres into 2 key spinal tracts with discrete yet overlapping functions. Within these tracts, the configuration of interneurons into localised circuits inhibit certain signals transmitted through fibres of varying myelination towards the brain, whilst allowing the excitability to remain in others when the scenario demands.
Take the edges of a 50p coin, having a distinct equilateral curve heptagon which can be discerned by the coordination of sensory information from the fingertips. The surface features of the coin face likewise stimulate the pathways of the brain which integrate the subtle features of an object, allowing for it to be identified without audible or visible recognition. This stereognosis precedes removal of the object into view, allowing its suspected identity to be confirmed, but becomes especially beneficial to individuals with visual impairment who rely greatly on this sense to read brail and so on. An adaptive compensation for loss of sight through heightening of tactile sensation is one example among many which the human nervous system is capable of.
The spinal cord tract responsible for transmitting these specialised sensations from the periphery of the body (the Dorsal Columnar Medial Lemniscal tract), which also provides us with the ability to sense fine touch and vibration, is hypothesised to have evolved after the spinal tract dedicated to crude tactile sensation, thermal sensation and pain (the Spinothalamic tract). Homeostasis and survival are the first functions of the nervous system in all species, and consequently, the almost endless adaptivity provided by the human brain and nervous system have allowed for the fruition of both survival and the range of high precision senses through which we comprehend our physical world. The cumulative development of these senses, such as our ability to determine the identity of an object through touch alone, has thus come to inform what makes us human.
Sensing & Sensibility…
Although the natural history of human life is echoed in the lengthy development of our nervous system, from its foetal to infant and adult form, it can be said in the wake of technology today that we are seldom exposed to the diverse ecology of materials and textures we evolved to detect, interpret and manipulate, by handling, shaping or otherwise learning to avoid if hazardous. We vie now for abstract information, at times broadcast at a global scale, and which can shape our thoughts, working lives and perceptions of the world at large. This is perhaps the inevitable conclusion of our being the most social species on Earth. As a result of what can be an excessive strain on our decision-making capacity, our attention is redirected increasingly to deal with this mass of information (2), often leaving the cognitive processes which would otherwise be dedicated to the sensation of our immediate world and situation, quite overwhelmed.
The nervous system has come frequently to forgo the sensation of other living things which exist beyond the infrastructure we have created. This itself may impinge our capacity to engage in our own emotions, and consequently, the emotions of others.
Within clinical aspects of anatomy and medicine this prompts some reflection: Do we humanise the cadaver of a donor beyond basic respect? How does the clinician approach a patient with empathy in a sustainable fashion, and whilst remaining objective? What potentiates the reciprocation of this empathy by a patient?
At a rudimentary level, we can reasonably say that the opportunity to communicate both shared and unshared experiences can foster trust in such scenarios. Our consideration of the first and foremost infrastructure in human life, the neuroanatomy on which our experiences and interactions depend, would necessarily seem to be conducive to this shared understanding.
Dexterity Within & Dexterity Without
The combined development of the upper limbs into highly dextrous and sensible (in the archaic sense of the word) appendages, and also of the lower limbs into weight-bearing and bipedal structures e.g., have had immense influence on the evolution of the axial skeleton (the skull, vertebrae and rib cage) and on the manner in which the nervous system maintains vital functions throughout the body. Sensation has also been pivotal to the expansion of the human brain, both in its processing and connections. Due to the ever increasing benefits of integrating select yet diverse sensory experiences, the course of vertebrate and mammalian evolution have been shaped by sensation, providing a foundation for the refined actions we use to respond to what we sense, whether in conscious reply to a question or as the typically unnoticed and unconscious reflexes which maintain the circulation of vital fluids.
That we are upright for example, has required a more complex array of mechanisms to maintain blood flow to our metabolically demanding brains, with all the conscious sensations we process, whether cutting wood, playing an instrument, writing, typing, driving and so on. Circulatory reflexes which maintain pressure and flow evolved initially at a lower centre of gravity in quadrupedal mammals, at their origin being more akin to the morphology of dogs, including those domesticated and selectively bred into various (and at times regrettably unhealthy) shapes and sizes.
Our nervous system however, being wired for bipedalism, and an upright centre of gravity, is calibrated to sense changes in blood pressure more acutely, and adapts the diameter of blood vessels more locally in order to accommodate blood flow to the brain and vital organs. Alterations to blood pressure e.g. are closely monitored and adjusted, especially in situations where the plane of gravity changes rapidly. Here the nervous system must prevent an average of ~800 ml of blood from pooling in the abdominal cavity and lower limbs; first by sensing the loss of pressure in the major veins and arteries adjacent to and above the heart, then initiating reflexes to correct the disparity in pressure and flow. Failure of this reflex is typically due to dysfunctions of the autonomic nervous system responsible for the reflex, leading to faintness brought on by low blood pressure and dysreflexias of the nerve impulses that elicit a complete cycle of blood in and out of the heart (normal sinus rhythm).
The brain, heart, kidneys and lungs are privileged over other organs, both in states of acute change to blood pressure and emergency (such as haemorrhagic loss of blood). Blood pressure is monitored internally (at the level of the neck) via sensory organs within the carotid (artery) sinus, operating in tandem with chemosensory changes in O2, CO2 and H+ (hydrogen ion = pH) levels in the carotid sinus of the same artery. These signals are largely conveyed via the vagus and glossopharyngeal nerves into the brainstem, not via the spinal cord, thus sensation from the viscera is not always the remit of the spinal cord, but in many instances is the responsibility of the 12 cranial nerves which project directly from the brainstem into the viscera of the head and all tissues beneath these.
If at all, how do the sensory and motoric functions of the spinal nerve tracts differ in their relation of what we experience, and how our bodies understand the world around us? Or the interoception of bodily tissues within us?
In this series, the architecture of nerves as the mechanism by which human beings are drawn towards advantage and away from danger will be explored, as well as the control that chemical and neuronal structures have in maintaining vital functions in everyday human life, and on how the known signs and symptoms of neuropathy are caused by abnormal alterations to these.
Frostell A, Hakim R, Thelin EP, Mattsson P, Svensson M. A Review of the Segmental Diameter of the Healthy Human Spinal Cord. Front Neurol. 2016;7:238. Published 2016 Dec 23. doi:10.3389/fneur.2016.00238
Wiehler A, Branzoli F, Adanyeguh I, Mochel F, Pessiglione M. A neuro-metabolic account of why daylong cognitive work alters the control of economic decisions. Curr Biol. 2022;32(16):3564-3575.e5. doi:10.1016/j.cub.2022.07.010
If you have enjoyed reading this article please consider subscribing to Lucida Visionis:
Or sharing its work with others: